ישנם בערך חמישה מסלולים בביולוגיה התפתחותית.
חזרה על השיעור הקודם:
Wnt:
הרצפטור זה frizzled הוא חוצה את הממברנה 7
פעמים ישנו גם קו רצפטור LRP ולשניהם אין שום פעילות
קטליטית הם רצפטורים שכאשרwnt נקשר יש גיוס של הקומפלקס (DVL GSK3 בטא וaxin-) לממברנה , DVL אותו GSK3 בטא שעושה בדרך כלל פוספורילציה על בטא catenin על מנת לסמן אותו
לדגרדציה עושה פוספורילציה על LRP ואז הפוספטים האלה מהווים
מגנט לaxin וכתוצאה מכך הקומפלקס לא נוצר ולכן ה- בטא catenin עובר אקומולציה בציטופלזמה נכנס לגרעין ושם הוא מחליף רפרסור
שיושב על TCF (פקטור שעתוק שיושב על הדנ"א) ,
אקטיבציה של שעתוק.

BMP:
מנגנון פעולה שונה.
הרצפטורים יש להם פעילות אנזימטית, האזור התוך תאי הוא בעל פעילות של סרין טארונין
קינאז ,ל- Type
2יש פעילות קונסטיטוטיבית ול- Type 1 יש פעילות אינאקטיבית,
כאשר יש דימריזציה Type 2 עושה פוספורילציה לעצמו
ול Type 1 ומאקטב אותו, עקב כך מתרחשת פוספורילציה על smad למינהם שהם פקטורי שעתוק
הם בצורה אינאקטיבית בציטופלזמה ובאיקטוב שלהם עם משמשים כפקטורי שעתוק בגרעין.

RTK=Receptor Tyrosine Kinase
משפחה של רצפטורים בעלי פעילות קטליטית באזור התוך תאי, פוספורילציה
על חומצה אמינית טירוזין.
Fgf:
ישנם 4 רצפטורים, ישנם הרבה splice variants אותו גן מייצר כמה רצפטורים של FGF. ישנם 20 ליגנדים של FGF. הרצפטור חוצה את הממברנה פעם אחת, באזור הלוץ תאי ישנם 3 דומיינים שנקראים אימונוגלובולינים דומיין (Ig domain) הדומיינים האלה קשורים לליגנדים, האזור התוך תאי הינו אזור אינאקטיבי זהו האזור של הקינאז, צריך לאקטב אותו על מנת לבצע פוספורילציה.

כאשר ישנו לנו ליגנד הרעיון של קישור הליגנד הוא שהוא (הליגנד) יוצר דימר, לוקחים שני רצפטורים ויוצרים בניהם דימר, הדימריזציה הזו בין שני הרצפטורים היא שמאקטבת את פעילות הקינאז, ולכן עכשיו כאשר הרצפטור בעל פעילות קטליטית הוא יכול לעשות אוטופוספורילציה -שם פוספט על עצמו. זה מאוד עקרוני במסלול הזה. למעשה הפוספטים האלה מהווים אתר עיגון לשני סוגים של חלבונים, לאנזימים ולאדפטורים. הפקטורים האלה שעוברים גיוס על גבי הרצפטור הם לא פעילים והם עוברים אקטיבציה ע"י פוספורילציה בזכות הרצפטור.

בתוך התא מבחינים בשלושה מסלולים שונים שהאקטיבציה של הרצפטור הזה
יכולה להוביל (לכן בעל תפקיד קרדינלי בסרטן) .
רואים בציור ליגנד שקושר שני רצפטורים לדימר. המסלול הצהוב שאותו
מתחיל PLC מוביל לעלייה בסידן וישנם שניים נוספים כחול
וירוק. אנחנו נתמקד בכחול- יצירת בדימר הביאה לאוטופוספורילציה שהיא הביאה לגיוס
של פקטורים כמו FRS2 חסר פעילות קטליטית, פקטור זו עובר פוספורילציה
ע"י הרצפטור מה שגורם לאקטיבציה שלו. ישנם שרשרת של חלבונים חלקם אנזימים
חלקם אדפטורים כאשר היעד זה לאקטב חלבון שנקרא RAS . ras נמצא בשני מצבים במצב
אקטיבי ובמצב אינאקטיבי.
כאשר GTP קשור ל- ras הוא נמצא במצב פעיל, כאשר GDP קשור לras הוא נמצא במצב אינאקטיבי. המעבר הזה מתווך ע"י חלבונים שונים. למשל GEF ( (Guanine nucleotide Exchange Factor החלבון הזה רואה את ה- GDP מעיך אותו ושם במקומו GTP. כאשר אנו רוצים לעשות ל ras אינאקטיבציה יש לעשות הידרוליזה לGTP ולהפוך אותו ל- GDP. ההידרוליזה הזאת יכולה להעשות את זה ע"י ras עצמו שיש לו פעילות של GTPase, אבל את הפעילות הזו צריך שמישהו יאקטב ואותה מאקטב GAP (GTPase Activating Protein). העבר ממצב אקטיבי ללא אקטיבי מתווך ע"י חלבונים.

למעשה sfp2 הוא פוספאטאז שמוריד
פוספטים על גבי הרצפטור שלפוספטים האלה נקשרים GAP – יוצר
משוב חיובי, הגברת סיגנל. SOS עושה אקטיבציה ל-RAS . RAS נמצא קשור לממברנה, ברגע
שהוא מאוקטב ישנה שרשרת אקטיבציה, כל החלבונים בשרשרת הם קינאזות, כל חלבון שעובר
פוספורילציה נהיה פעיל כאשר המטרה היא לעשות פוספורילציה ל MAPK-
- עובר טרנסלוקציה לגרעין
ושם הוא משפעל פקטורי שעתוק ע"י פוספורילציה. הערה SFP2 הוא GAP שרוצה לעקב את RAS.
במסלול הזה לא מכירים אנטגוניסטים חיצוניים לתא, כל האנטגוניסטים
שמכירים הם בתוך התא. לדוג' אנטגוניסט טרנסממברנלי שהוא משפיע על הרצפטור ישירות.
ישנו sprout שגם
הוא משפיע על המסלול.
Shh:
מסלול נחקר מאוד,עם החורים הכי גדולים. Shh שמור באבולוציה אבל יש לו
מורכבות מסויימת שעדיין לא מבינים אותה. במסלול הזה הוא יתייחס גם לעכברים וגם
לדרוזופילה. בדרוזופילה יש ליגנד אחד בעכברים ישנם שלושה ליגנדים- Shh, Ihh, Dhh (3 ליגנדים זה לא מורכבות כ"כ גדולה
ולכן זה קצת מוזר) . בדרוזופילה יש רצפטור אחד ובעכבר יש שני רצפטורים Ptc1, Ptc2. ישנו Smo שנמצא אחד בדרוזופוליה
אחד בעכבר. ישנם פקטורי שעתוק שנקראים Gli1-3 (3 בעכבר). בדרוזופילה זה
נקרא Ci קיים אחד בדרוזופילה. המורכבות עלתה אהל לא
באופן משמעותי.
המסלול המתואר מתאים
גם לדרוזופילה וגם לעכבר:
הרצפטור PTC חוצה את הממברנה 12 פעמים, ישנה מולקולה נוספת שנקראת SMO שחוצה את הממברנה 7 פעמים והיא מעבירה את הסיגנל. PTC כרצפטור מעכב את SMO . כאשר אין ליגנד בסביבה אין סיגנל כי PTC עושה איהיביצה לSMO. כאשר אין ליגנד ישנו קומפלקס שנקרא Cos2/Kif7 (אדפטור) אחד החלבונים החשובים זה FU, חלבון חשוב נוסף הוא Su(fu). כאשר אין סיגנל יש פקטור שעתוק שנקרא GliFL/Ci155 כלומר זה חלבון במשקל של 155 קילודלתון והוא במצב של Full length במצב שלם (חלבון צהוב). כאשר אין סיגנל הוא מגוייס לקומפלקס, עובר פוספורילציות מסוימות כאשר הן מסמנות אותו לעבור דגרדציה חלקית ע"י הפרוטאוזום ולבסוף אנחנו נשארים עם GliR/Ci75, והוא נכנס לגרעין יושב על הדנ"א ועושה עיכוב שעתוק. כאשר יש Shh (ליגנד) הוא נקשר לראצפטור Ptc כתוצאה מהקישור הזה האינהיביציה של SMO הולכת ויש גיוס של הקומפלקס שסימן מקודם את GliFL/Ci155 לדגרדציה את הממברנה וכתוצאה מכך הלא עובר - GliFL/Ci155 לא עובר פוספורילציה ולכן לא מתבקע ע"י הפרוטאוזום, נשאר שלם. (הציור הצבע של החלבון מתשנה מצהוב לירוק וזאת משום שחושבים שהוא משנה את הקונפורמציה שלו עדיין לא יודעים מה קורה שם ממש) החלבון הזה (GliFL/Ci155) נכנס לגרעין ומאפשר שעתוק.

בדרוזופילה Ptcנמצא בממברנה אבל SMO לא נמצא בממברנה הוא נמצא באנדוזום. ישנה הפרדה בין Ptc ל- Smo ,אבל כאשר Shh נקשר לרצפטור מאיזושהי סיבה smo עובר לממברנה והוא מאוקטב הוא עובר פוספורילציה. בזמן ש-Shh נקשר לרצפטור קורים שני דברים, אחד מהם זה טרנסלוקציה של smo מהאנדוזום לממברנה ואקטיבציה של smo עצמו, עדיין לא יודעים איך זה קורה שם.
כיצד קורית האינאקטיבציה? זה נכון גם לעכברים וגם לדרוזופילה (עם הבדלים מסויימים) המנגנון אינהיביציה הוא לא מנגנון סטוייכומטרי כלומר אין צורך על כל Ptc אחד Smo על מנת לעשות את האינהיביציה, אין קישור של שתי המולקולות ולכן יש עיכוב. יש צורך במעט מולקולות של Ptc על מנת לעשות אינהיבציה להרבה מאוד מולקולות של Smo, לכן ייתכן כי זהו תהלי ך אנזימטי או שזהו תהליך של טרנספורטר. העדויות על עכשיו תומכות בתהליך של טרנספורטר-Ptc 12 פעמים חוצה את הממברנה והוא מבחינה הומולוגית דומה לטרנספורטים שונים ולכן הוא יכול לייצר תעלה כזו שתאפשר למולקולות להיכנס ולצאת מהתא. בציור רואים מודל בו יש מעבר של חומרים דרך הרצפטור שמעכבים את Smo וכאשר הליגנד נקשר הוא חוסם את המעבר ואין מעבר של המעכב ממקום אחד לשני ואז Smo אקטיבי (אין מודל מוכח לזה, זה מה שמשערים).

Primary cilia:
להרבה תאים יש
מיקרווילי , מיקרווילי זה מין בלוטה שיוצאת מהתא, האורך שלה הוא מאוד קטן. בתמונה
רואים הרבה מיקרווילי ורואים בליטה אחת ארוכה שזוהי בעצם ה- cilia ,וזה נקראה primary cilia כי בדרך כלל לתאים אנימיילים לרוב התאים יש
סילייה אחת כזו, יש לה כל מיני תפקידים.היום יודעים שהיא דרושה לסיגנלים לתוך התא
ביונקים. בתמונה רואים את הפריימרי סילייה
באדום. (השוטון בתא זרע לדוגמא זה פריימרי סילייה).

המבנה המולקולרי של הפריימרי סילייה:
רואים שיש ליבה ,יש
את ה- Axoneme שהוא
מורכב ממיקרוטובול, הממברנה ממש צמודה למיקרוטובול. רואים גם שהתנועה לתוך הסילייה
ומתוך הסילייה לתא היא לא דיפוזיילית, היא תנועה אקטיבית. יש תנועה לתוך הסילייה –
Anterograde transport ומתוך הסילייה לתא – Retrograde transport. התנועה היא אקטיבית יש צורך
"במכוניות" בין אם זה חלבון ממברנלי או חלבון תאי. יש טרנספורט אקטיבי,
יש צורך בהשקעת אנרגיה על מנת להביא לסילייה או מהסילייה.

נחזור ל- Shh- ביונקים גם רואים מצב
שבו ה- smo נמצא
באנדוזום אבל אחד הדברים החשובים ביונקים זה איזשהו הפרדה בין Ptc ל- Smo כאשר Ptc נמצא על גבי הסילייה על הממברנה ו mso נמצא על הממברנה מחוץ
לסילייה ( יש פעמים שהוא נמצא על האנדוזום ואז יש צורך להביא אותו לממברנה). כאשר
אין Shh , Ptc לא מאפשר ל smo להגיע לפריימרי סילייה. ל-Gli יש שתי אפשרויות או שהוא עובר דגרדציה מלאה או שהוא עובר ע"י
הקומפלקס בצורה אקטיבית לפריימרי סילייה,ושם הוא עובר מודיפיקציה, עובר
פוספורילציה , ביקוע ויצירת הרפרשור שמגיע לגרעין.
כאשר יש לנו Shh אשר נקשר ל- ptc ישנה טרנסלוקציה כפולה ptc עובר א הפריימרי
סילייה הולך אל הממברנה ו smo לעומת זאת עובר לפריימרי
סילייה שם הוא הופך להיות אקטיבי והוא עושה את כל פעילות הסיגנלנילג שלו שתביא
בסוף לאקטיבציה של Gli (את זה לא רואים בדרוזופילה, לא יודעים למה זה)
יודעים שאם מורידים את הפריימרי סילייה דופקים את המסלול של Shh. העניין הזה מדליק נורה
של מורפוג'ין, Shh הוא אחד המורפוגניים
הקלאסיים בתהליכים ההתפתחותיים.
התא שמייצר את הסיגנל: מיהו Shh? זהו חלבון שהוא פוליפפטיד בעל שני דומיינים, דומיין אחד 19 קילודלתון והשני 26 קילו דלתון. הליגנד עצמו הוא הדומיין של ה- 19 דילודלתון, ה-26 זה דומיין שדומה לאינטהין ( תופעה שבה protein splicing חלבון ארוך שבאמצע יש מקטע שיודע לצאת ולחבר את החלקים האחרים וליצור חלבון חדש) הדבר הזה דומה לאינטהין הוא לא אינטהין. יש לו פעילות פרוטאוליטית, פרוטאז שעושה ביקוע ואז מחבר כולסטרול (נעשה ב- ER) במצב זה הוא לא פעיל רק אחרי שמוסיפים קבוצה ליפידית בקצה ה-N טרמינלי וואז זה הליגנד פעיל. מי שעושה את זה הוא אנזים. לליגנד הפעיל יש גם כולסטרול וגם קבוצה שומנית. Shh נוצר ב- ER, Ski זה אותו חלבון שיודע לעשות תוספת של קבוצה שומנית, את הכולסטרול מוסיף הדומיין, ואז מה-ER הוא מגיע לממברנה רק שיש גם קבוצות של כולסטרול וליפיד ולכן הוא לא ישתחרר בקלות מהתא, ובשביל זה יש חלבון שנקרא Disp והוא יוצר קומפלקס גדול שהוא יכול לעבור ולהגיע רחוק למרחקים גדולים. בלי Disp לא נוכל לקבל את זה (לא יודעים איך). תופעה שאיננה קיימת בטבע באופן טבעי: יוצרים מוטציה שבה הכולסטרול לא מוסף ואז מקבלים משהו פעיל שיוצא החוצה אבל הוא לא יצור מולטימר ולכן הפעילות שלו תהיה לטווח קצר.

Notch:
במסלול הזה בשונה מהמסלולים השונים הליגנד מחובר לממברנה ואיננו משתחרר . הרצפטור נקראה Notch יכולים להיות כמה 1,2,3.. וכו' הליגנד גם הוא חלבון טרנסממברנלי יש לו כמה שמות ( Delta לדוגמא) לליגנדים, כדי שתא אחד ידבר עם תא שני הם חייבם להיות קרובים ולכן הם לעולם לא יהיה long rang . האינטראקציה בין הליגנד לרצפטור חייבת להיות בין שני תאים קרובים. הליגנד נקשר לרצפטור וע"י זה נוצרת קסקדה של אירועים, זה מתחיל בשני ביקועים, אחד נעשה ע"י Tace שנמצא על גבי הממברנה (קורה בגלל החיבור של הליגנד והרצפטור) הביקוע הזה חותך מעל הממברנה ומשחרר את האזור האקסטרהסלולרי שנשאר צמוד לליגנד ורק אז מתרחש הביקוע הבא ע"י קומפלקס ענק שנקרא גמא secretase, קומפלקס זה עושה ביקוע בתוך הממברנה, הביקוע הזה משחרר את האזור התוך תאי, החלק התוך תאי הזה הוא פקטור שעתוק, הוא משתחרר נכנס לתא ועושה אקטיבציה של שעתוק ע"י החלפת הרפרסור. החלק של הרפרסור שקשור לליגנד עובר אנדוציטוזה לתא ודגרדציה יחד עם הליגנד.
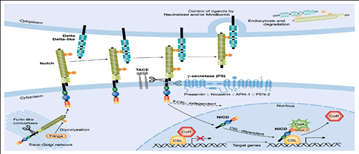
איך נוצר הרצפטור? ב- ER שהחלבון הזה נוצר הוא
נוצר כפוליפפטיד אחד,הוא חוצה את הממברנה פעם אחת. ואז ישנו אנזים שנראה o-fut הוא מוסחף קבוצות סוכר של פוקוז על גבי החלבון. הוא יכול להוסיף
את זה בכל מיני מקומות, האנזים הזה משמש גם כשפרון באופן בלתי תלוי והם מגיעים
ביחד לגולג'י, שם ישנו סרין פרוטאז שנקרא פורין furin והוא עושה ביקוע וחותך את
הרצפטור לשניים אבל הם לא משתחררים הם נשארים כבולים, בלי הביקוע הזה הרצפטור
איננו אקטיבי. ואז בגולג'י מגיע Fringe והוא מוסיף קבוצות סוכר
נוספות לסוכר דבר אשר משנה את האיפיניות לליגנד ולכן תפקידו מאוד חשוב! אותו
רצפטור מגיע לממברנה. הליגנד נוצר בממברנה בצורה לא אקטיבית, כדי לאקטב אותו הוא
צריך לעבור אנדוציטוזה ישנה פעילות קטליטית שמורידה את כל האזור האנטרסלולרי ואז
מחזירים אותן לממברנה בצורה האקטיבית שלו. ורק אז האינטרציה בין הרצפטור לליגנד
יכולה להתרחש.
לסיכום: המסלולים האלה שונים אחד מהשני במנגנון, בחלבונים, בשיטה
וכו.. אבל לא כל מסלול הוא באופן עצמאי, הם מדברים אחד עם השני! זה חשוב בביולוגיה
התפתחותית. בכל רקמה ובכל תא הם מדברים אחרת תלוי בקונטקסט.
Feedback loops: אותו מסלול/תהליך משפיע על עצמו מעצמו. יכול
להיות העצמה או דיכוי של סיגנל. זה נכון לגבי כל המסלולים. ישנו משוב חיובי
(העצמה) ומשוב שלילי (דיכוי).
•
Bottle neck of
the pathway: צוואר בקבוק של המסלול. בתוך ביולוג התפתחותי רוצים לדעת כיצד מסלול
מסויים משפיע על תהליך מסויים, מחפשים את צוואר הבקבוק שכאשר נפגע בו נפגע בכל
המסלולץ לדוגמא ב wnt צוואר הבקבוק הוא בטא catinin (הוא אחד ויחיד) במקרה של
Shh זה Smo. וזה מביא להבחנה
בין Loss of
function שזה ניסוי שבו לוקחים גן ועושים לו החסרה ורואים מה זה עושה, ישנם
ניסויים שהם Gain of
function לוקחים פקטור שרוצים לדעת מה הוא עושה ורואים כיד הוא פועל
במקומות שונים.